
Salt plante


Salt planter eller Halofyter (fra gammel græsk ἅλς hals , "salt" og φυτόν phyton , "plante") danner en økologisk afgrænset gruppe blandt de højere planter , der er tilpasset forøgede niveauer af let opløselige salte i deres placering og kan gengive under disse betingelser.
Saltplanter koloniserer saltrige steder stort set uanset bredde i tørre til oversvømmede levesteder, ofte nær havet og på saltsøer . De endnu lidt undersøgte mekanismer, hvormed planter kan tilpasse sig ekstreme miljøforhold og forblive fotosyntetisk aktive, er meget forskellige. Nogle saltplanter (de obligatoriske halofytter ) fremmes i deres vækst ved moderat saltindhold, andre saltplanter behøver ikke salt til deres vitale aktivitet. De trives meget bedre på saltfri jord, men der er de ringere end andre planters konkurrence .
Definitioner
Der er flere definitioner af udtrykket salt plante eller halofyt. Et kriterium er for eksempel, at planterne kan vokse på salt jord med mere end 0,5 procent salt i den tørre jordvægt. Selvom denne værdi er let at bestemme, er salt- eller ionkoncentrationen i jordvandet vigtigere for planten. Definitionen af Jennings (1976), som også ofte bruges, beskriver halofytter som den naturlige flora på saltsteder . Allerede Warming (1909), der definerede udtrykket Halophyt første gang, skrev: skal være til stede, før en halofytisk vegetation fører til en vis mængde opløselige salte; typen af salt synes ikke at have betydning. Mann et al. (1983) beskrev lokaliteterne således: "[...] indeholder saltvand med et osmotisk tryk på over 3,3 bar" , hvilket svarer til en koncentration på 70 m M monovalente salte.
Planter, der ikke overlever på saltsteder, selvom konkurrence er udelukket, kaldes ofte glykofytter i tysktalende lande . Dette udtryk stammer fra ordet "ferskvand" og kan kun findes på det tyske sprog. Mere passende, men mindre hyppigt anvendte navne for det samme udtryk er halofobe planter eller simpelthen ikke-halofytter .
Salt-tolerante organismer omtales generelt som halofile , når de ikke længere er i stand til at eksistere i fravær af salt på grund af den evolutionære saltresistens. Denne saltresistens har udviklet sig uafhængigt flere gange. Derfor er der ingen saltplanter med monofyletisk afstamning, selvom de i nogle planteslægter og - familier ofte forekommer.
Blandt de højere planter findes halofytter kun blandt angiospermerne. Der er omkring 1.500 kendte arter af halofytter . Familier med mange repræsentanter er det Chenopodiaceer , aizoaceae , Frankeniaceae , blyrod-familien og blandt mangrover den mangrovetræ-familien , Lythraceae , Avicennioideae i akantus familien , Combretaceae og Myrsinaceae . Der er også halofytter blandt de søde og sure græsser og Juncaceae .
Der er 54 halofytter i Tyskland. Der er også nogle overgangsformer og mellemarter, der sjældent udvikler halotolerance eller betragtes som hybrider af arter af littoral ( Ammophila arenaria ) og indlandet ( Calamagrostis epigeios ). Eksistensen af sådanne særlige kystformer af ellers udbredte halofytiske eller sandlevende græsser er det bedste bevis på, at disse må have taget deres nuværende placering i besiddelse i meget lang tid og skabt de former, der svarer til det.
Klassificering efter saltafhængighed
Obligatoriske halofytter
De obligatoriske halofytter ( obligatoriske = obligatoriske ), også kaldet Eu -halofytter, er bundet til deres salte miljø. Uden en bestemt koncentration af salt som grundlag for liv, ville disse planter ikke kunne trives og spire, da de stort set har tilpasset sig de ekstreme forhold ved denne miljøfaktor. Den tolerance vifte af de obligatoriske Halofyter til salt er derfor meget store, så de kan eksistere selv med konstant oversvømmelse med havvand. De bedst kendte indenlandske slægter er samphire ( Salicornia ), siv ( Juncus ), Salzmelden ( Suaeda ) og silt græsser ( Spartina ).
Fakultative halofytter

De fakultative halofytter ( fakultativ = valgfri) har evnen til at vokse på saltsteder, men er ikke bundet til dem. Repræsentanter som gås ukrudt ( Potentilla anserina ), strand aster ( Aster tripolium ), strand plantain ( Plantago maritima ) og strand mugwort ( Artemisia maritima ) kan også forekomme i maritime områder. Imidlertid opnår de kun deres optimale vitale funktioner på jord, der overvejende er saltfri eller kun har et let saltindhold. Da de i stigende grad støder på konkurrence fra andre planter i disse områder, har disse saltplanter ofte en ulempe i forhold til de ferskvandsplanter, der findes der. Disse er bedre tilpasset deres miljø og formerer sig hurtigere. Fakultative halofytter har et mere begrænset toleranceområde for jordens saltholdighed end obligatoriske halofytter.
Ligegyldige halofytter
Såkaldte placeringsindifferente halofytter danner en overgangsform til ferskvandsplanterne og findes normalt kun i saltfrie områder. Deres toleranceområde er relativt lille, men de kan stadig klare saltjord, der har en lavere koncentration. I disse tilfælde ændres habitus for den tilsvarende plante på forskellige måder og afviger fra det grundlæggende udseende. Repræsentanter for denne gruppe er: rød svingel ( Festuca rubra litoralis ), hvid struds græs ( Agrostis stolonifera ), tudse dunhammer ( Juncus bufonius ), lav ranunkel ( Ranunculus repens ) og stenurt ( Sedum ).
Klassificering efter saltets påvirkningstype
Salt kan virke på planten på forskellige måder, ifølge hvilke halofytter dybest set kan beskrives som luftthalen ( aerohalin ), vandhalin ( hydrohalin ) eller jordbaseret halin . De to sidstnævnte kategorier hænger sammen, og derfor bruges samlebetegnelsen hydroterrestrisk halin ofte.
Air thalenes

Brydebølger og sprøjtning på havene fører gennem en dispersionsproces ( bulk til partikelomdannelse ) til frigivelse af små dråber ( havspray ) i luften. Betydelige dele af den transporteres opad af turbulensen af det marine grænselag og kan delvis tørre ud. Den aerosol, der dannes ved en sådan surf -atomisering , som i fællesskab betegnes havsalt -aerosol , har en effekt på planter i områder nær og langt fra havet på grund af den høje koncentration af saltluft. Mange air-thalin-arter er også vandhalin, så der er en tættere forbindelse mellem de to kategorier. De strengt aerohaline arter lever som en overgang fra det maritime til det terrestriske område i fjerntliggende omgivelser til havet og optager salt næsten udelukkende gennem bladoverfladen. Saltindholdet i bladene på vindsiden kan være op til ti gange højere end bladene på den samme plante i slipstrømmen. Den gule hornvalmue ( Glaucium flavum ) er ufølsom over for saltstøv og stænkvand, men tolererer ikke salt i jorden.
I nærheden af fordampningsbassiner, dvs. visse indre saltsøer, der regelmæssigt tørrer ud i perioder med tørke og efterlader en saltørken ( ørkendannelse ), er der også mange arter af saltplanter. Saltet i luften kan spores tilbage til sådanne fordampningsprocesser i saltvandet og absorberes af floraen, der lever der fra det aerobe miljø.
Vandhalin
De marine arter lever i nærheden af både saltvand og brakvand og kan findes fra tidevandszonen via flodmundinger ind i det indre saltvand . Hydrohaline planter er alle de arter, der enten er helt eller halvvandede , dvs. har deres livscenter i eller i nærheden af vandet. Hvis jorden er tør og sandet, i snævrere forstand på strande og klitter, har de arenikoløse halofytter der normalt tilpasset sig deres miljø med et yderligere, dybere rodsystem. På den mudrede, ofte oversvømmede undergrund, som sikrer direkte vandoptagelse, har planter med mindre, men også stærkere, rødder, der ikke kan vaskes væk, en tendens til at dukke op. Nogle lukkede plantedannelser har åbenbart specialiseret sig i udtyndet havvand ved floddeltaerne og flodmundinger som levesteder, hvorimod andre lever i en udtalt sandgapflora på saltkoncentrerede søer og indlandshav såsom Det Døde Hav . Saltindholdet i thalassohalin -steder varierer over et bredt område og kan svare til saltvand i saltvand (3,5%) op til saltindholdet i en mættet natriumchloridopløsning (30%). Overgangsformerne til de jordbaserede og aerohaline saltplanter danner også områder med mangfoldighed fjernt fra saltvand, som har lagt sig på normal jord eller stenbrokker (fortrinsvis Sedum -arter) over vandstanden, der normalt når ved højvande og normalt kun nås af vandet salt spray .
Terrestrisk halin
Terrestriske haliner er alle terrestriske arter, der har specialiseret sig i saltvand i indlandet . De optager kun salt gennem jorden. Salt planter, der ikke vises på stranden omfatter for eksempel klippe bynke ( Artemisia rupestris ), slids endivie bynke ( Artemisia laciniata ) eller jord ridning græs ( Calamagrostis epigeios ). Også planter, der forekommer i ørken- og steppeområder, såsom arter af slægten Atriplex, skal forstås som former fjernt fra vandet, som fortrinsvis kan findes under lokaliserings-ligegyldige halofytter .
Klassificering efter jordens saltholdighed
Klassificeringen efter saltindholdet i jorden er klassificeret ved hjælp af data i promille (1 ‰ svarer til 1 g / L). Da mange terrestriske halofytter også er vandhalin, gælder klassificeringsskemaet for både kategorier og også for geologi og jordvidenskab .
- Oligohalin (græske oligoer - små ) planter tåler en saltkoncentration på 0,5 til 5 ‰ og har et meget lavt toleranceinterval med hensyn til salt. De fleste marine og terrestriske arter tæller i denne rækkefølge.
- Mesohaline (græsk meso - midten , midten ) er navnene på planter, der er placeret i områder med 5 til 18 ‰ saltindhold. Fællesskaber i denne økologiske gruppe findes i nærheden af saltmyrer eller søer med medium saltholdighed.
- Polyhaliner (græsk poly - flere , meget ) har et saltindhold på op til 30 ‰ i deres miljø. Det er planter, der lever på usædvanligt salte søer.
Virkninger af salt på planten
Saltets virkninger er synlige på tre niveauer. I tilfælde af ikke-halofytter fører dette til skade, mens halofytterne tilpasses af forskellige mekanismer.
- Osmotiske virkninger gør det svært at optage vand fra det salte medium ( fysiologisk tørhed ).
- Overskuddet af natrium fører til en ion -ubalance, da de vitale ioner kalium, calcium og nitrogen ikke længere kan absorberes tilstrækkeligt af planter, der ikke har tilpasset sig. Der er en natriuminduceret kaliummangel.
- Saltioner har også specifikke virkninger på forskellige metaboliske områder. Eksempler er inhibering af proteinmetabolisme, en stigning i aktiviteten af pentosefosfatcyklussen og et fald i glykolyse .
Blandt jordsalte har natriumchlorid den største toksicitet og de fleste steder også den største andel; Dette efterfølges af calciumchlorid , magnesiumchlorid , natrium og magnesiumsulfat afhængigt af styrken af den giftige virkning .
Placeringer
Bortset fra polarområderne i Arktis og Antarktis er halofytter almindelige på alle kontinenter. De kan findes i europæiske havområder såvel som i fugtige , tropiske regnskove eller tørre saltstepper, på strandene på fjerntliggende øer i Oceanien , i et alpint område på op til 3.100 meter høje saltsøer og saltkilder eller i midt i ørkenen ( Sahara ) på natriumrige sand. Selvom de er repræsenteret i alle ikke-polære klimazoner, er de tropiske og tempererede breddegrader de vigtigste udbredelsesområder.
På det biogeografiske niveau har størstedelen af de kendte arter et rumligt begrænset udbredelsesområde. Planter spredt over hele verden, såkaldte kosmopolitere , findes kun sporadisk og udelukkende under obligatoriske halofytter, såsom saltbusken .

Primære placeringer
Alle naturlige og næsten naturlige levesteder i Europa er beskyttet af sjældenheden af individuelle saltplantearter. Disse naturlige levesteder, der ikke er skabt eller forårsaget af mennesker, kaldes primære steder. De har normalt en stor befolkningstæthed . Den biologiske mangfoldighed er ofte relativt lav på grund af den ekstreme placering.
De to vigtigste kystnære levesteder for halofytterne er saltmyrer, der forekommer i tempererede, og mangroveskove, der forekommer i tropiske og subtropiske klimaer. Næsten alle saltplanter, selvom de er delvist resistente over for xerofytisk dehydrering, er afhængige af en lokal vandkilde for konstant saltfortynding.
Saltmose levested

I kystlinjen områder af tempererede klimazone, der er mudrede moser områder på flade kyster inden for den midterste oversvømmelse linje , der er oversvømmet ved højere vandstand. Sådanne salte områder, kaldet saltmyrer , er levested for mange blomstrende planter, der er tilpasset disse ekstreme forhold. Afsætningen af saltmasserne finder sted afhængigt af højden over middelfloden og dermed på saltindholdet og er opdelt i forskellige zoner alt efter vegetationen .
I Tyskland er saltmoserne stort set beskyttet, for eksempel i Niedersachsen Nationalpark Vadehavet .
Mangroveskovs levested

Økosystemet for de stedsegrønne mangrover findes i tidevandsområdet for flade, tropiske kystområder . Især i de varme områder omkring ækvator , z. Artfattige tidevandsskove, nogle mere end 30 m høje. Mangrovetræer er et af de få træer med en udtalt saltmodstand. De overjordiske dele af rodsystemet i mange mangrovearter er slående (f.eks. Stilterødder i Rhizophoreae , pneumatoforer i Avicennia osv.), Som forsyner ilt med det underjordiske rodsystem i det anoksiske sediment og øger træernes stabilitet. I hvilket omfang deponering af sedimenter i området med mangroverødderne stabiliserer kysten, kan diskuteres.
Flere steder
I Centraleuropa er saltlande i landet ganske sjældne. Kendte eksempler er saltmose omkring Kyffhäuser-bjergene i Thüringen , Pannlake i Hollerlands naturreservat i Bremen, Luchwiesen i Brandenburg eller Salzlacken i Seewinkel øst for Neusiedl-søen . To faktorer er vigtige for forekomsten af saltlokationer ud for havets kyst: Der er saltaflejringer under jorden og i hvert fald i nogle sæsoner er fordampningen højere end nedbøren, så saltene når jordoverfladen og ikke vaskes ud. Det er netop på sådanne steder, at der ikke kun forekommer natriumchlorid, men også magnesium , carbonater og sulfater .
Desuden findes halofytter sjældnere i bjergene, hvorved salte fanget i klippen eller saltkilder og søer i højder er grundlaget for dette lidt usædvanlige levested. I løbet af dagen tilbyder den ofte stenede undergrund varmeabsorbering , så planterne ikke dør af kulde om natten. Også typisk for klipper salt-tolerante Felsbesiedler hvordan kan Samphire , hav raket , hav roer og strand Lilac forekomme. De er i stand til at forankre deres rødder dybt i klippen for ikke at blive skyllet væk af stormvande.
.jpg)
Saltplanter optager store områder i de tørre (tørre) områder af jorden, som modtager mindst nok nedbør til, at saltene kan koncentrere sig om overfladen med det stigende grundvand på grund af fordampning. Saltplanterne på tørre steder kaldes xerohalophytes. I det følgende præsenteres nogle områder med xerofytisk vegetation som eksempler.
I Ebro -bassinet (Spanien) mødes det kontinentale klima med afløbsløse saltsøer. En række endemiske gypsophytes, for eksempel Lygeum spartum, vokser på gips-rige jorde . I Australien indtager Chenopodiacean semi-desert ( Saltbush ) store områder i den sydlige del af kontinentet. Arter af slægterne Atriplex og Maireana vokser på de salte jordarter (kun omkring 0,1% chlorid) , mens ikke-halofytter (akacier osv.) Vokser på de sandede områder. De Maireana buske kan leve op til 300 år. I Iran , Afghanistan og store områder i Centralasien er jordbunden salt. I det centrale Afghanistan forekommer der stadig halobiomer på 3.100 meter over havets overflade (Dasht-e-Nawor). De indre iranske bassiner, Kawire, er ofte ekstremt salte og absolut fri for vegetation. Typiske haloserier rige på Chenopodiacea forekommer i kantområderne. I de centralasiatiske ørkener forekommer halofytvegetation i fugtige fordybninger ( saltpander , shory) og omkring saltsøer . Den mest saltresistente saltplante i disse områder er den pudedannende Halocnemum strobilaceum (Chenopodiaceae). For Karakum -ørkenen er den halofytiske busk Haloxylon persicum ( White Saxaul , Chenopodiaceae) den karakteristiske plante. Saltlokationerne i Centralasien strækker sig over Det Kaspiske Hav til Europa. På den nordlige del af Krim-halvøen er der det dovne hav (Sivash), der tørrer om sommeren, og hvorfra salt blæses mod nord, hvor det forårsager sodafstivning ( solonation ) af jorden. Saltjordene i det sydlige Rusland kaldes Solontschake . I Great Basin i Nordamerika ved Great Salt Lake er der enorme saltområder. Den vigtigste art er Atriplex confertifolia (engelsk shadscale ), der dækker omkring 150.000 km² i USA. I Death Valley og San Joaquin Valley i Californien forekomme borax rige salt websteder, som især Atriplex hymenelytra , Suaeda Torreyana og Bassia koloniserede arter. Saltpander kan findes i de sydamerikanske pampas , f.eks. Laguna La Picaza i provinsen Santa Fe , hvor sodavæskning kan nå pH -værdier på 10, og hvor især distichlis græs vokser. I mere tørre områder forekommer kloridsulfatopbygning oftere, især i den vestlige del af pampaerne.
Sekundære placeringer
Ud over de primære steder er der sekundære, menneskeskabte saltområder på minedrift og industriområder samt langs trafikveje, dvs. dem, der kun blev skabt gennem menneskelig påvirkning. Floraen af sådanne levesteder tilhører ruderalvegetationen , da det er urteagtige arter, der ikke bruges til landbrug eller skovbrug. Den vigtigste lokaliseringsfaktor er det øgede saltindhold, som er forårsaget af forskellige handlinger, såsom saltning af vejene om vinteren; mekaniske forstyrrelser spiller kun en sekundær rolle. Sekundære levesteder repræsenterer ø -levesteder for halofytter og halobionter , så de også er af betydning for biogeografisk forskning. Undersøgelsen af afviklingen af overbelastning og resthavere af kaliumindustrien , der blev klassificeret som "øbjerge" i kulturlandskabet, har vist sig at være særlig interessant . Indtil nu er der fundet sjældne eller ikke -observerede arter, der tidligere kun var kendt fra primære steder, inde i landet. I forhold til befolkningsstørrelserne for mange truede halofytter fungerer de sekundære steder nu som vigtige tilflugtssteder.
Tilpasningsstrategier
Saltplanter har udviklet en række strategier for at kunne vokse, når de udsættes for salt. " En plantes evne til at holde en for stor tilførsel af salte i sit substrat væk fra protoplasmaet ved at regulere salt eller at udholde en øget osmotisk og ion-toksisk saltbelastning " omtales som saltresistens. Modstand er derfor det generiske udtryk, de to underudtryk er regulering og tolerance.
Saltresistens manifesterer sig i morfologiske og fysiologiske tilpasninger, som for det meste er gensidigt afhængige.
Saltregulering
afskærmning
Afskærmningen (engelsk eksklusion ) er strategien for ikke at absorbere saltioner i planten eller ikke lade dem komme ind i følsomt (ungt, voksende) væv.
Afskærmning i roden
På grund af eksklusionsprincippet, også kendt som saltfiltrering, er optagelsen af saltioner via rødderne begrænset. Som med alle planter forhindres mangrovefamiliens ukontrollerede apoplastiske vandoptagelse af den kaspiske strimmel . Ionkanalerne i cellemembranen er meget mere selektive end ikke-halofyters, så næsten ingen natrium og chlorid absorberes, men vigtige ioner som kalium absorberes. Den xylem saft inden roden er lav i salt, i modsætning til jorden. Når der i kanten af mangroveskove voksede busk Conocarpus erectus ( combretaceae ) i xylemsaften på grenene var der kun 17,6 mol m -3 chlorid og 7,5 mol m -3 (målt natrium i forhold til 465 mol m -3 chlorid og 362 mol m -3 natrium i havvand).
Afskærmning i scion
Hos nogle arter, for eksempel Prosopis fracta , en busk i tørre saltområder , bevares især natriumioner i plantens basale dele, såsom rødder og stamme ( natriumretention ). Det samme gælder for den Andel græs ( Annelgræs peisonis ). I græsset Diplechna fusca bevares natrium- og chloridioner i bladskeden . I tilfælde af mangroverne, som denne strategi ville være passende på grund af bagagerummet, kunne ingen sådan mekanisme observeres.
Ved at bevare saltioner i de basale og også ældre dele af planten holdes ionindholdet i de unge skudafsnit og også i de fysiologisk aktive blade lavt.

B = blærecelle, C = neglebånd,
Ch = kloroplast, E = epidermale celler,
M = mesofylceller, P = plasmodesmata,
S = stamcelle, V = vakuoler
eliminering
Eliminering er strategien for at udskille saltioner, der allerede er absorberet af planten. Der er mange forskellige strategier her. Udskillelsesmekanismerne er en særlig effektiv, men også energiforbrugende, metode til direkte udskillelse af det indtagne salt og dermed holde cellens saltværdi næsten konstant.
Afsaltet hår
Blødningshår eller blærehår er særlig almindelige i gåsefodfamilien (Chenopodiaceae). Disse er specialiserede hår ( trichomer ) på bladoverfladen, som ioner aktivt transporteres til. Håret dør, brister, ionerne vaskes af og fjernes dermed fra planten. Blærehår er for det meste tocellede, de består af en blærecelle, der er næsten fuldstændigt fyldt med vakuolens cellesaftrum, og som sidder oven på en vesikel, plasma, mitokondrier og kloroplastrige stamceller. Sidstnævnte forbinder blærecellen med det underliggende bladvæv gennem talrige plasmodesmata og indtager også en meget metabolisk og fysiologisk aktiv position for at muliggøre transport af natriumchlorid med den efterfølgende akkumulering. Med energiforbrug ( ATP ) transporteres opløste salte via stamcellen ind i blærecellens vakuol og akkumuleres der. Ved et forudbestemt bristepunkt på skaftet knækker håret, når det har absorberet nok salt og falder til jorden. Hvis denne mekanisme mislykkes, brister håret eller fjernes ved nedbør med vaskning og afvaskning.
Nogle arter, såsom Atriplex halimus eller purslane kilen ( Halimione portulacoides ) kan udskille mere end 80% af de ioner, de absorberer gennem hårene på deres vabler.
Afsaltningskirtler

Mange halofytter udskiller salte gennem særlige kirtler på bladoverfladen. I modsætning til udskillelse og sekretion betegnes saltudskillelse som rekretion , dvs. H. stofferne udskilles af planten i den form, de blev indtaget i, og de tjener ikke længere noget særligt formål efter udskillelsen.
Saltkirtlerne i de enkelte halofytter viser meget forskellige strukturelle og funktionelle træk. En af de enkleste former findes i slægt af silt græs ( Spartina ). I deres blade og stilke, de normalt kun bestå af en pore- fyldt kutikula og en subkutikulær rum med underlaget og ventil celler.

I almindelig havlavendel ( Limonium vulgare ), som har en meget kompleks struktur af saltkirtlerne, mistænker man eksocytose som en ekstra mulighed for materialetransport. De akkumulerede saltvesikler ( exosomer ) smelter sammen med sekretionscellernes membran og kommer til overfladen gennem porer. Dette kan medføre, at der dannes synlige saltkrystaller på bladets yderside. Antallet og fordelingen af saltkirtlerne i bladet er også forskelligt. Mens lavendel har op til 3000 kirtler pr. Cm², viser nelliken ( Armeria maritima ) kun 590 kirtler og mælkeblomst ( Glaux maritima ) omkring 800 kirtler pr. Cm².
Natrium- og chloridioner udskilles specifikt, mens kalium bevares i planten, så kalium / natriumforholdet i planten forbliver højt. Mængden af salt, der udskilles af kirtlerne, kan nå meget høje værdier, for eksempel kan Diplachne fusca udskille fem gange mængden af salt, der absorberes på samme tid, græsset Spartina alternifolia omkring det halve. Mangrove aegialitis annulata udskiller omkring 38% af saltmængden i unge blade på 12 timer. Energibehovet for saltudskillelse er betydeligt. Ved Tamarix kræves omkring 20 til 24 mol ATP pr. Mol udskilt natriumchlorid.
Afgivelse af plantedele
Rosetplanter akkumulerer saltioner i vævene på de ældste blade, indtil toksicitetsgrænsen er nået. Inden disse dør, fjerner planten de genanvendelige næringsstoffer ( nitrogen ) fra dem og kasserer dem derefter med det lagrede salt. De efterfølgende yngre blade overtager funktionen. Velkendte Central europæiske repræsentanter er strand vejbred ( Plantago maritima ), strand trefork ( strand-trehage ) og strand aster ( Aster tripolium ).
Omranslokation
Retranslokation forstås som returtransport af ioner fra bladene via floemet tilbage til rødderne, hvor ionerne frigives tilbage i det omgivende medium. Denne mekanisme er blevet bevist for det søde græs Pappophorum pappiferum , som kan udskille omkring 35% af natrium i skuddet via rødderne inden for 48 timer. Denne mekanisme diskuteres også for mangrover, da blade af mange arter kan leve op til 18 måneder med den samme saltholdighed.
Succulens

_02.jpg)
Succulens er en strategi til fortynding af det indtagne salt; det forekommer primært med chloridhalofytter. Vand absorberes også med ionerne og opbevares i de store vakuoler . Dette forhindrer en for høj intracellulær saltkoncentration. Afhængigt af det saftige organ skelnes der mellem blad succulenter som sod ( Suaeda ), ostedug ( Spergularia ) og nogle mangrove -arter og stammesukkulenter såsom samphire . Succulens forekommer særlig hyppigt i gåsefodfamilien. I bladets saftige mangrover Rhizophora -mangel ( Rhizophora -familien ), Laguncularia racemosa og Conocarpus erectus (begge vingede familie ) er der en høj korrelation mellem chloridindholdet baseret på bladoverfladen og vandindholdet baseret på bladoverfladen; denne sammenhæng forekommer ikke i saltvandsmangrover.
De fleste sukkulenter er kendetegnet ved kødfulde, hævede skud og roddele, der er blevet omdannet til vandopbevaringsorganer af lag med flere lag. Bladene, hvor antallet og størrelsen af parenkymceller øges, har et reduceret overfladeareal og er ofte konisk i stammen for at begrænse sved. Med akkumulering af vand forårsager alle fortyndingsmekanismer en stigning i volumenet af opløsningsrummet og et fald i saltionkoncentrationen.
Alternativet til opbevaring i cellevakuolen er fordelingen af det overskydende salt i hele planten, hvilket i første omgang resulterer i en reduktion i saltkoncentrationen. I tilfælde af den årlige samphire eller saltrus , er vegetationscyklussen allerede forbi, når saltkoncentrationen bliver dødelig . Den saltoverbelastede plante bliver brun til rød - et generelt symptom på stress - og dør til sidst.
Intracellulær opdeling
De forskellige fordeling af de absorberede ioner til forskellige cellekomponenter ( cellelokaler ) er placeret som en tilpasningsmekanisme mellem regulering og tolerance. Selv planter, der opbevarer salte, kan ikke modstå nogen ionkoncentrationer i cytoplasmaet , da de fleste af deres enzymer også er følsomme over for salt. Derfor er de fleste saltioner fanget i vakuolen . Dette er især mærkbart med sukkulenterne med deres store vakuoler, men opdelingen er ikke begrænset til succulenterne. Cytoplasmaet og chloroplasterne indeholder derfor kun en relativt lav koncentration af saltioner. Iontransporten ind i vakuolerne foregår gennem membran -ATPaser . Den resulterende forskel i det osmotiske potentiale mellem vakuol og cytoplasma afbalanceres med kompatible opløste stoffer (se nedenfor).
tolerance
Larcher definerede salttolerance som følger: Salttolerance er den protoplasmiske bestanddel af resistens, nemlig evnen til at tolerere skiftet i ionforholdet, der opstår under saltstress og de toksiske og osmotiske virkninger forbundet med den øgede ionkoncentration, afhængigt af plantearterne , vævstype og vitalitetsstatus. Denne salttolerance skyldes flere biokemiske mekanismer.
Membransammensætning
Saltplanter har en anden sammensætning af deres cellemembraner . En højere andel mættede fedtsyrer reducerer membranernes fluiditet og reducerer dermed saltionernes udslip fra vakuolen. Dette er en vigtig forudsætning for at opretholde ovennævnte opdeling.
Ionpumpemekanismer
For at opdele saltionerne skal saltplanterne have transportmekanismer.
Den ophobning af natrium- og chloridioner i vacuolen er afhængig af en pH -gradient, der er bygget op af tonoplasten ATPase. PH -gradienten mellem cytoplasma og vakuol muliggør derefter en H + / Na + antiport, gennem hvilken natrium kommer ind i vakuolen. I strandbanan ( Plantago maritima ) for eksempel dannes disse H + / Na + antiportere kun, når der er saltstress, men ikke når der mangler salt i vækstmediet.
Kompatible opløste stoffer
Akkumuleringen af de uorganiske saltioner i vakuolen skaber en forskel i koncentration til cytoplasmaet. For at afbalancere dette osmotiske potentiale syntetiserer saltplanter organiske forbindelser, som de akkumulerer i cytoplasma og chloroplaster. Disse kompatible, dvs. ikke-toksiske, opløselige osmotika kaldes kompatible stoffer eller, med deres engelske navn, kompatible opløste stoffer . Koncentrationen af de kompatible stoffer afhænger af saltbelastningens omfang.
Ofte kompatible stoffer er aminosyrer ( prolin ), kvaternære ammoniumforbindelser såsom glycinbetain , sukker samt acykliske ( sorbitol , mannitol ) og cykliske sukkeralkoholer (såkaldte cyclitter, for eksempel pinitol ). Sukkeralkoholer forekommer især i mangrover. Nogle havalger som Dunaliella akkumulerer glycerol , andre som Chlorella saccharose.

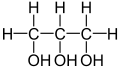
Proline forekommer ofte i planter under stress
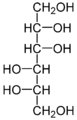
Mannitol , en sukkeralkohol, der ofte findes i mangrover som et kompatibelt stof

Sorbitol bruges f.eks. I Plantago maritima siden
Algen Dunaliella akkumulerer glycerol under saltstress




{kind=link}
{kind=link}
Transgen tobak har vist, at kompatible stoffer faktisk øger salttolerancen . Tobak er normalt følsom over for salt. Som et resultat af den genetiske manipulation af det gen for enzymet mannitol-1-phosphat-dehydrogenase, de modificerede planter akkumulerede mannitol og, som et resultat, voksede signifikant bedre end vildtype, når de udsættes for salt.
Nogle planter opbevarer store mængder organiske stoffer i vakuolen for at opbygge det osmotiske potentiale, der er nødvendigt for vandoptagelse, og er derfor ikke afhængige af optagelsen af saltioner. Disse stoffer omfatter oxalat , malat , aspartat (asparat) og saccharose . Disse øgede andele af organiske stoffer er særligt mærkbare i tilfælde af løvfisk , siltgræs og havlavendel .
Fotosyntetiske mekanismer
I nogle planter aktiveres fotosyntesens CAM -vej under saltstress . Dette gør det muligt for planterne at fotosyntese med meget lidt fordampning af vand. Dette reducerer plantens vandoptagelse fra jorden og dermed også saltoptagelsen. Dette reducerer plantens saltbelastning. Disse valgfrie CAM -planter omfatter Mesembryanthemum crystallinum , Mesembryanthemum nodiflorum , Aptenia cordifolia og Carpobrotus edulis .
Som et C 4- anlæg har strandbananen ( Plantago maritima ) også en vandbesparende fotosyntesetype.
| Tilpasningsmekanismer i halofytter i de tyske saltmyrer | ||||||
| Halofytter (udvalg) | Justeringsmekanismer | |||||
| akkumulering | Succulens | Blærehår | Bladfældning | Saltkirtler | undtagelse | |
| Andel ( Puccinellia maritima ) | x | - | - | - | - | - |
| Dansk spoonweed ( Cochlearia danica ) | x | x | - | x | - | - |
| Strand nellike ( Armeria maritima ) | - | x | - | x | x | x |
| Samphire ( Salicornia europaea ) | x | x | - | - | - | ? |
| Brine rush ( Juncus gerardii ) | x | - | - | x | - | - |
| Salt alarm ( Atriplex halimus ) | x | - | x | - | - | - |
| Siltgræs ( Spartina ) | x | - | - | - | x | x |
| Strand aster ( Aster tripolium ) | x | x | - | x | - | - |
| Strandkrus ( Artemisia maritima ) | x | - | - | - | - | x |
| Strandtrident ( Triglochin maritima ) | x | x | - | x | - | - |
| Havlavendel ( Limonium ) | x | - | - | - | x | x |
| Strandode ( Suaeda maritima ) | x | x | - | - | - | - |
| Strandbanan ( Plantago maritima ) | x | x | - | x | - | x |
Fremhævet med grønt : særlig stærkt
Funktioner og brug
Halofytter såsom samfire eller spadestik tilhører pionerplanterne og kan bidrage til sedimentering og dannelse af saltmoser i havområder. Mens rødderne holder jorden på plads, sikrer den øverste del af planten, at vandbevægelsen er rolig. Saltet og andre sedimenter, der føres af havvandet , aflejres mellem rødderne og individuelle dele af planten. De synker derefter ned i jorden. En langvarig gentagelse af denne proces kan føre til, at bunden fortsat stiger og stiger over vandstanden. Sådanne undersøgelser giver igen den mindre saltresistente flora et levebrød. Denne tilsmudsningseffekt bruges lejlighedsvis ved at skabe "halofyt-senge". Bølge- og vinddæmpende effekt er også velegnet i hele verden som en naturlig buffer og beskyttende funktion mod stormfloder og tsunamier ( genplantning af mangroverne i Vietnam, Thailand og Indien).
En anden nyttig fordel er forebyggelse af slid på strande og kyster. De brede rod løbere på nogle planter (f.eks fælles strand græs (sand-hjælme) , også strand rug ( Elymus arenarius )) anker selv over lange afstande i jorden og styrke den. Dette vil forebyggende fjernelse af materiale, der forhindres af vandet. På denne måde kan vandindtrængning forhindres eller endda stoppes på truede øer eller bugter.
Det statlige forskningscenter Desert Research Center (DRC) i Kairo leder efter metoder, som egyptiske ørkenregioner kan gøres brugbare. De Maryout Eksperimentel Station dækker omkring fire hektar og er DRC største forsøgsstation. Det har studeret adfærden hos dyr og afgrøder under ekstreme forhold siden 1968. Hovedforskningsfokus er på, hvor godt geder, får, kaniner og kameler tåler foder fra planter, der vokser i jorden, selv ved høje saltkoncentrationer. Indtil videre er husdyrs reaktioner på to typer saltplanter som ernæringskilde blevet testet, hvor forskere sammenlignede vækst, frugtbarhed og kødkvalitet med forsøgsdyr med normale foderstoffer. Atriplex halimus ( saltbuske ) og Acacia saligna ( saltacacia ) begrænsede kun væksten i forsøget, hvilket kan kompenseres for med fedtholdig mad. I dag tjener de som fødekilde på særligt tørre steder. Halofytter bruges også til at stabilisere marker uden at begrænse væksten af afgrøder. I mellemtiden er det brugbare areal i ørkener blevet stærkt forøget gennem blandet dyrkning med saltplanter .
Mange saltmose planter bruges også som mad i køkkenerne i de nordlige lande. Spiselige saltplanter (såsom strandbanan , samphire som havasparges eller strandtricorn som rørkål) giver retterne en delikat, salt til peberagtig smag og tjener som en naturlig kilde til jod for kroppen .
Desuden blev asken fra brændende halofytter tidligere brugt til en lang række produktionsgrener, såsom glas og sæbe, men dette er ikke længere tilladt på grund af den faldende artsbestand.
Referencer
- ↑ a b c d e f Marianne Popp: Saltresistens i urteagtige halofytter og mangrover . Progress in Botany, bind 56, Springer Verlag Berlin 1995, s. 416-429.
- ↑ a b H. Bothe: Saltresistens i planter , 1976.
- ↑ Biologisk leksikon. Bind 7 . Spektrum Akademischer Verlag, Heidelberg 2001, s 33,. ISBN 3-8274-0332-4 -
- ↑ H. Walter, S.-W. Breckle: Jordens økologi. Bind 1: Grundlæggende . G. Fischer, Stuttgart, 2. udgave 1991, s. 102-109, ISBN 3-437-20454-8 .
- ^ Karlheinz Kreeb: Planter på saltsteder . Naturvidenskab 61 , 1974, s. 337-343
- ↑ Walter Larcher: Planteøkofysiologi . Eugen Ulmer, Stuttgart, 4. udgave 1995, s.316.
- ↑ a b Christiane von den Berg: Salzwiese habitat - halofili eller : plantetolerancemekanismer mod saltstress , status: februar 2005 (adgang: 9. april 2006).
- ↑ a b Dieter Schlee: Økologisk biokemi . Gustav Fischer, Jena, Stuttgart 1992, s. 170-187, ISBN 3-334-60393-8 .
- ↑ a b Universitetet i Osnabrück: Planter på saltvandssteder (halofytter) ( Memento fra 27. november 2005 i internetarkivet ) (adgang: 7. april 2006).
- ↑ Walter Larcher: Planteøkofysiologi . Eugen Ulmer, Stuttgart, 4. udgave 1995, s. 319.
- ↑ MC Tarczynski, RG Jensen, HJ Bohnert (1993), Science 259, pp. 508-510.
- ↑ Hilmar Liebsch; Michael Thekat: Salt - et problem for landbruget I: Quarks Sendungsarchiv. Quarks & Co, udsendelse: 23. august 2005 (adgang 21. april 2006).
litteratur
- Centraleuropæisk saltvegetation
- Ernst Albert Arndt: Mellem klitten og havbunden. Dyr og planter i Østersøregionen . Leipzig / Jena / Berlin 1969.
- A. Gerhardt: Kystens planter. Videnskab i klasseværelsets biologi. Aulis, Köln 1982,5, s. 164-174. ISSN 0342-5487
- HJ Janssen: Vadehavets fare . BUND oplysninger. Bind 20. Freiburg 1983.
- F. Jantzen: Planter ved havet. Landbuch, Hannover 1968, 1978, 1987. ISBN 3-7842-0363-9
- BP Kremer: Planter ved vores kyster. Stuttgart 1977, 1999. ISBN 3-440-07734-9
- Richard Pott: Color Atlas North Sea Coast og North Sea Islands . Ulmer, Stuttgart 1995. ISBN 3-8001-3350-4 (god, omfattende illustreret beskrivelse af vegetationen på det tyske Nordsø)
- G. Quendens: Strand og kyst . BLV, München 1984, 1988.
- HE Reineck (red.): Watt. Kramer, Frankfurt 1978, 1982. ISBN 3-7829-1067-2
- M. Thies: Vadehavets biologi . Aulis, Köln 1985. ISBN 3-7614-0795-5
- Saltvegetation på verdensplan
- Georg Grabherr: Color Atlas of Earth's Ecosystems. Naturlige, næsten naturlige og kunstige landøkosystemer fra et geobotanisk synspunkt. Ulmer, Stuttgart 1997. ISBN 3-8001-3489-6 (kort oversigt)
- Heinrich Walter, Siegmar-W. Breckle: Jordens økologi . 4 bind. Gustav Fischer / Elsevier, Stuttgart / München 1991–2003. ISBN 3-437-20297-9 (detaljeret beskrivelse af jordens vegetation og dens økofysiologi, herunder halofytter)
- fysiologi
- R. Albert: Halofytter . I: H. Kinzel: Planteøkologi og mineralsk metabolisme. Ulmer, Stuttgart 1982, s. 33-204. ISBN 3-8001-3427-6
- R. Albert, G. Pfunder, G. Hertenberger et al.: Fysiotype tilgang til forståelse af halofytter og xerofytter . I: S.-W. Breckle, B. Schweizer, U. Arndt (Hrsg.): Resultater af verdensomspændende økologisk forskning . Heimbach, Stuttgart 2000, s. 69-87.
- W. Baumeister, W. Ernst: Mineralsk metabolisme og plantevækst. Fischer, Stuttgart 1978. ISBN 3-437-30271-X
- Marianne Popp: Saltresistens i urteagtige halofytter og mangrover . I: Fremskridt i botanik. Bind 56. Springer, Berlin 56.1995, s. 416-429. ISSN 0340-4773
- IA Ungar: Økofysiologi af vaskulære halofytter . CRC Press, Boca Raton 1991. ISBN 0-8493-6217-2
- Y. Waisel: Halophyters biologi. Fysiologisk økologi. Redigeret af TT Kozlowski. New York / London 1972. ISBN 0-12-730850-4
Weblinks
- Halofytter i botanik ( erindring fra 29. december 2011 i internetarkivet ) (oversigt over yderligere weblinks)
- Saltmose levested
- Liste over halofytter
- Naturpark omkring Kyffhäuser -bjergene (PDF -fil; 4,94 MB)
- Tyske nationalparker i Nordsøens vadehav